

bat365正版唯一官网
《Cell Host & Microbe》和《Science Signaling》封面文章报道系统所游富平课题组在细…
|
|
天然免疫免疫从上世纪发现模式识别受体在现在一直学界研究的热点,而细胞核内的天然免疫识别和调节机制还不为人所知。2019年3月19日,bat365官网登录入口系统生物医学研究所游富平研究组研究成果以“HIPK2 is necessary for type I interferon-mediated antiviral immunity”为题,作为封面文章发表于Science子刊Science signaling,HIPK2作为蛋白激酶在细胞核内磷酸化ELF4并启动抗病毒基因转录(图1)。2019年9月11日,游富平课题组在Cell 子刊《Cell Host & Microbe》期刊上发表题为“The Nuclear Matrix Protein SAFA Surveils Viral RNA and Facilitates Immunity by Activating Antiviral Enhancers and Super-enhancers”的论文,揭示了SAFA作为一个新型的细胞核内病毒dsRNA监测器,连接染色质重塑和抗病毒天然免疫反应(图2)。
| 图1. Science Signaling 封面文章 图2. Cell Host & Microbe备选封面 |

病原生物入侵机体后,产生的病原相关分子模式(pathogen-associated molecular pattern,PAMP)被一系列的模式识别受体(pattern recognition receptors,PRRs)识别,最终诱导各种炎症因子、趋化因子以及干扰素(Interferon,IFN)的产生,发挥抗感染功能。模式识别受体是识别外来危险信号的感受器,并且传递信号给细胞内的效应分子,从而调动细胞进行免疫应答。首先被发现的模式识别受体是Toll 和Toll 样受体(Toll like receptors),Jules A Hoffmann和Bruce Beutle 因为他们在这方面的工作获得了2011年的诺贝尔生理医学奖。所有的TLRs 都无一例外的定位于细胞质膜和内体。接下来科学家发现了在细胞胞浆内的模式识别受体包括:RIG-I、MDA5、cGAS、DDX41、Caspase-4/5/11和NLRs等,能够识别细胞胞浆内的病原生物。然而在病毒感染过程中会在细胞核内产生大量的DNA和RNA,细胞核内的模式识别过程还有待深入研究。已知IFI16能够识别细胞核内的病毒DNA并且激活STING相关信号通路。就在上个月,南开大学曹雪涛院士团队发现hnRNPA2B1识别细胞核内的病毒DNA并激活抗病毒天然免疫反应。但是细胞核内是否存在识别病毒RNA的模式识别受体还有待揭示。
病毒RNA,尤其是双链RNA(double-stranded RNA,dsRNA)是一种重要的PAMP,病毒感染宿主细胞产生的dsRNA可被TLRs家族的TLR3或RLRs家族的MDA5(melanoma differentiation association gene 5)和RIG-I(retinoic acid-inducible gene I)识别。TLRs家族一般定位在细胞膜或胞内体上,能识别细胞外或经过吞噬进入细胞的PAMP。RLRs家族定位在细胞质,识别细胞质内的核酸组分。研究显示,多种病毒感染会在宿主细胞核中产生dsRNA1-2,但细胞是否存在监测机制识别细胞核中病毒dsRNA目前还未有报道,本研究首次揭示了细胞核内病毒RNA感受器的存在。
另一个重要的科学问题是在感染条件下,宿主细胞如何调节染色质的开放性从而打开以及关闭一系列基因的表达,从而使机体清楚感染并维持免疫稳态和健康。本研究首先通过全基因组功能性筛选发现SAFA能够协同病毒激活抗病毒天然免疫反应。SAFA(Scaffold Attachment Factor A),又叫HnRNPU(heterogeneous ribonucleoprotein U,和曹雪涛院士团队发现的hnRNPA2B1是同家族分子),作为最早被发现的核脚手架蛋白,对于核内部结构的维持起重要作用。研究表明SAFA参与染色质空间结构的调控,SAFA功能异常与X染色体失活、扩张型心肌病、肿瘤等多种疾病相关。本研究发现SAFA在识别病毒RNA之后发生活化,并调节染色质的开放性以及免疫基因增强子和超级增强子的活性。
该研究显示多种病毒感染宿主细胞后,在细胞核中产生dsRNA,被SAFA监测结合,引起SAFA发生寡聚化。SAFA同病毒RNA的结合是本研究亮点和关键点,研究人员运用生化手段,从各个层面和多个角度证明SAFA在病毒感染情况下同病毒dsRNA有很强的结合。并且通过RIP-seq (RNA Immunoprecipitation-sequencing,RNA免疫共沉淀测序)和TRIBE(targets of RNA-binding proteins identified by editing,基于核酸编辑的RNA结合蛋白鉴定技术) 编辑3(图2)等二代测序手段进一步肯定了SAFA和病毒RNA的结合。RIP-seq的结果显示SAFA倾向于结合HSV-1病毒在细胞核内产生的dsRNA。SAFA通过其C端的RGG结构域结合病毒RNA,这一结果也得到了近期研究的支持-RGG倾向于结合dsRNA,而不是ssRNA(single strand RNA,单链RNA)3。为了进一步确证SAFA能够和病毒的dsRNA相结合,研究人员构建了SAFA和ADAR1催化结构域的融合蛋白。ADAR1是一种RNA脱氨酶,能够识别dsRNA上的腺嘌呤并且对其进行脱氨,使得腺嘌呤变成次黄嘌呤。构建了ADAR1催化结构域和SAFA的融合蛋白的稳定表达细胞株之后,病毒感染该细胞,并把细胞中的RNA进行全基因组的RNA-seq分析,发现HSV-1病毒RNA发生了脱氨反应,由此进一步确认了SAFA能够同病毒dsRNA结合。
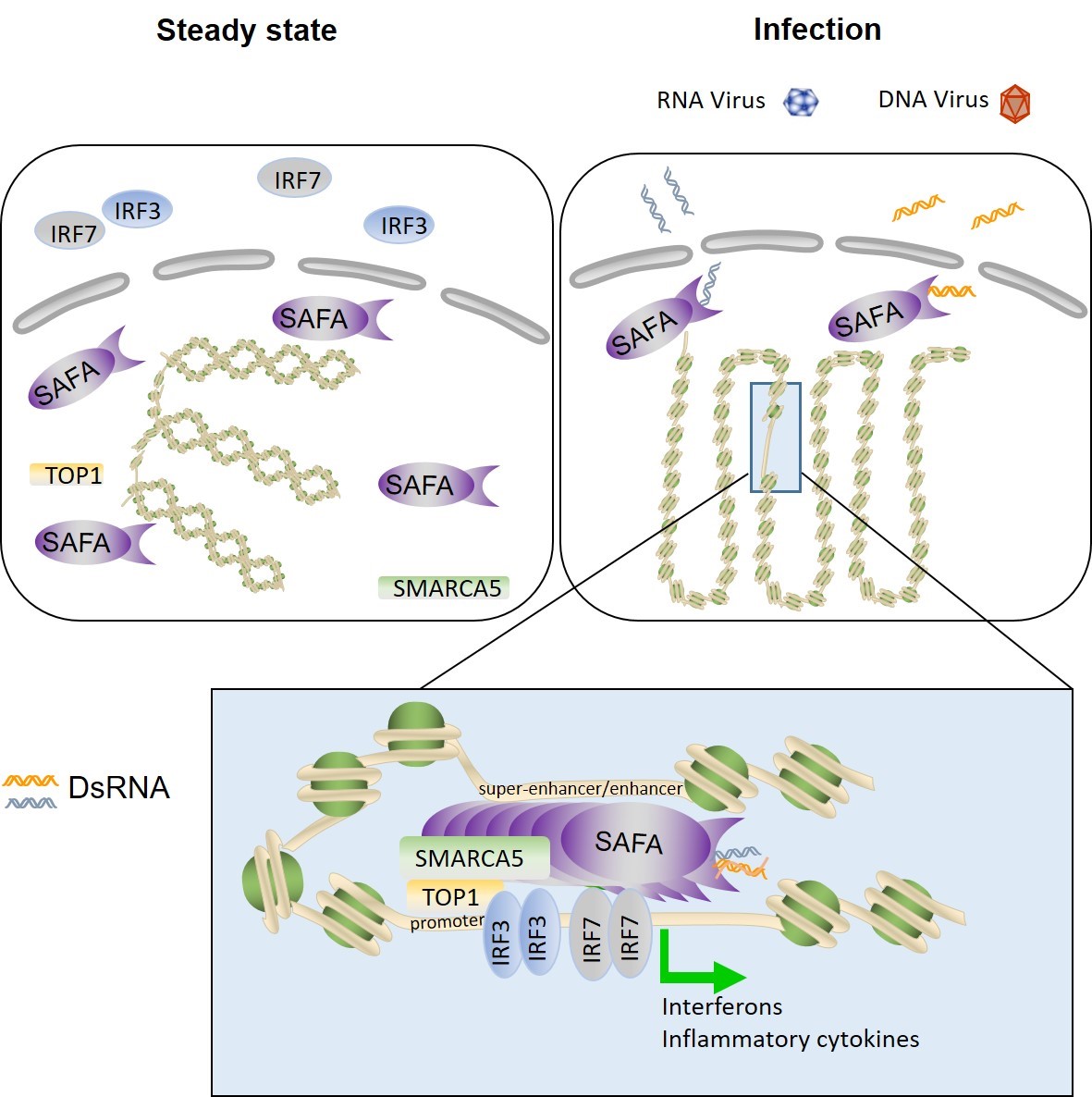
图3. SAFA识别细胞核内病毒RNA并调节抗病毒基因增强子活性
该研究首次鉴定了细胞核内病毒RNA的模式识别受体,并且SAFA在识别了病毒RNA之后通过调节染色质重构以及基因远端增强子的活性来调节抗感染基因的表达,在传统研究免疫信号通路激活转录因子的基础上,率先引入了天然免疫信号对于基因远端增强子的调节机制(图3)。因为在后生动物,包括小鼠和人类中,远端增强子对于基因转录的起始和维持可能比启动子区域扮演了更加重要的角色。所以,研究远端增强子和超级增强子对于免疫应答调控和免疫细胞命运决定,是十分有意义的,此部分工作应该只是一个小小的开始。而该研究中还有很多问题没有解决,包括:SAFA如何调控相关基因染色质和增强子的调节特异性;SAFA如何发生的多聚化以及其调节机制;在感染过程中增强子以及超级增强子的动态调节过程,以及对免疫记忆和免疫耐受的调节。
|
图4. Science Signaling 同期评论文章的模式图介绍HIPK2作用机制 |
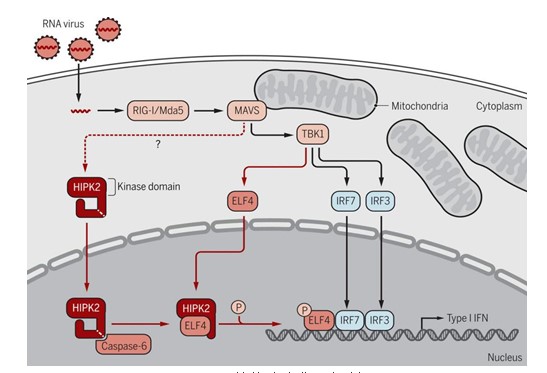
Science Signaling的文章中作者阐明了HIPK2在机体产生I型干扰素抗击RNA病毒感染中的关键作用。敲除HIPK2后巨噬细胞对水泡口炎病毒诱导产生的I型干扰素水平显著降低,敲除HIPK2的小鼠对于致死性剂量的水泡口炎病毒感染更加敏感。进一步的机制研究发现,水泡口炎病毒感染后,活性的Caspase酶切割HIPK2,释放出HIPK2的在N-末端片段并转移入细胞核,HIPK2入核片段与ELF4 蛋白结合并促使ELF4的第369位丝氨酸发生磷酸化,ELF4与IRF3、IRF7共同启动I型干扰素的转录,发挥抗RNA病毒作用。
Science signaling同期发表“Focus”评论文章,专门对本项研究进行了评述。文章指出同样具有酶活性的HIPK2和Caspase6很有可能作为多种自身免疫性疾病的治疗靶点,通过没活性调节干扰一型干扰素的表达来治疗相关疾病(图4)
bat365官网登录入口免疫系的2015级博士生操丽丽和刘胜德为Cell Host & Microbe共同第一作者,操丽丽同样是Science Signaling的第一作者,bat365官网登录入口系统所游富平研究员为通讯作者。研究得到国家自然科学基金和蛋白质机器重点专项等项目的经费资助。
参考文献:
1. Wang L, Wen M, Cao X. Nuclear hnRNPA2B1 initiates and amplifies the innate immune response to DNA viruses. Science. 2019 Aug 16;365(6454).
2. Kurilla, M.G., Piwnica-Worms, H., and Keene, J.D. (1982). Rapid and transient localization of the leader RNA of vesicular stomatitis virus in the nuclei of infected cells. Proc Natl Acad Sci U S A 79, 5240-5244.
3. Weber, F., Wagner, V., Rasmussen, S.B., Hartmann, R., and Paludan, S.R. (2006). Double-stranded RNA is produced by positive-strand RNA viruses and DNA viruses but not in detectable amounts by negative-strand RNA viruses. J Virol 80, 5059-5064.
4. Ozdilek, B.A., Thompson, V.F., Ahmed, N.S., White, C.I., Batey, R.T., and Schwartz, J.C. (2017). Intrinsically disordered RGG/RG domains mediate degenerate specificity in RNA binding. Nucleic Acids Res 45, 7984-7996.
5. McMahon, A.C., Rahman, R., Jin, H., Shen, J.L., Fieldsend, A., Luo, W., and Rosbash, M. (2016). TRIBE: Hijacking an RNA-Editing Enzyme to Identify Cell-Specific Targets of RNA-Binding Proteins. Cell 165, 742-753.